1) 1 o 2 FS.
2) Cadena transportadora deelectrones.
3) ATP-sintetasa

En los cloroplastos hay un transporte no cíclico, con fotólisis del agua y reducción del NADP+, además de síntesis de ATP, en el que intervine el FS II y el FS I y uno cíclico, como el de las bacterias, en el que no se rompe la molécula de agua y, por tanto no se reduce el NADP+, sólo se forma ATP:
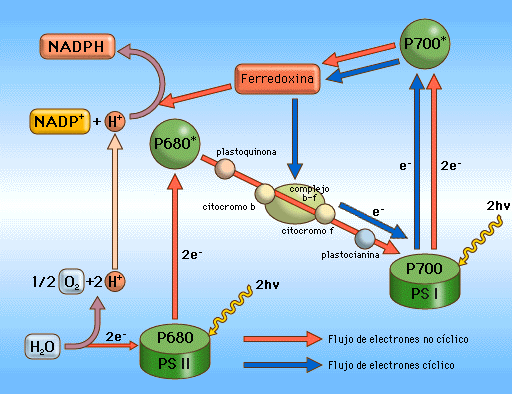


1. Fotofosforilación no cíclica:
a) Fotólisis del agua

b) Transporte de los electrones del agua a través de la cadena transportadora de electrones:
Agua------P680 (FS II), que recupera los electrones perdidos al incidir la luz sobre él (el pigmento diana del FS)
Los e* (electrones excitados) del P680--------------------Plastoquinona (estado energético menor)
PQ (plastoquinona)-----------------Complejo de citocromos b-f (E.E. menor)
cit b-f----------------------------------PC (plastocianina) (E.E. menor)
PC-------------------------------------P700 (FS I), que recupera el electrón que ha perdido al incidir sobre él la luz.
Los e* del P700-------------------Fd (ferredoxina)
Fd------------------------------------Complejo NADP-Reductasa, que junto con H+ del estroma, produce NADPH en el estroma:
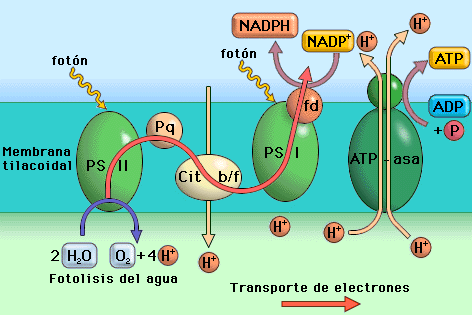
A nivel del cit b-f la energía liberada en la caída energética de los electrones se emplea en tranportar contragradiente H+ desde el estroma al tilacoide, contribuyendo a aumentar más la concentración de H+ dentro del tilacoide: quimioósmosis.
c) Reducción del NADP+ por la NADP-reductasa, complejo enzimático que está en la cara del estroma de la membrana tilacoidal, formándose NADPH.
d) Fotofosforilación del ADP, con formación de ATP, cuando los H+, que están más concentrados dentro del tilacoide salen al estroma ( a favor de gradiente), por la ATP-sintetasa.
2. Fotofosforilación cíclica
a) Transporte de electrones. La luz incide sobre el FS I, de modo que sus e*------------Fd
Fd----------cit b-f, que cuando cede los electrones a la PQ libera la energía suficiente para bombear contragradienre H+ al interior del tilacoide: quimioósmosis.
PQ---------------PC-----------------FS I
B) Fotofosforilación del ADP.
Cuando salen los H+, a favor de gradiente, desde el interior del tilacoide al estroma, lo hacen a través de la ATP-sintetasa, aportando la energía necesaria para fabricar ATP en el lado del estroma.
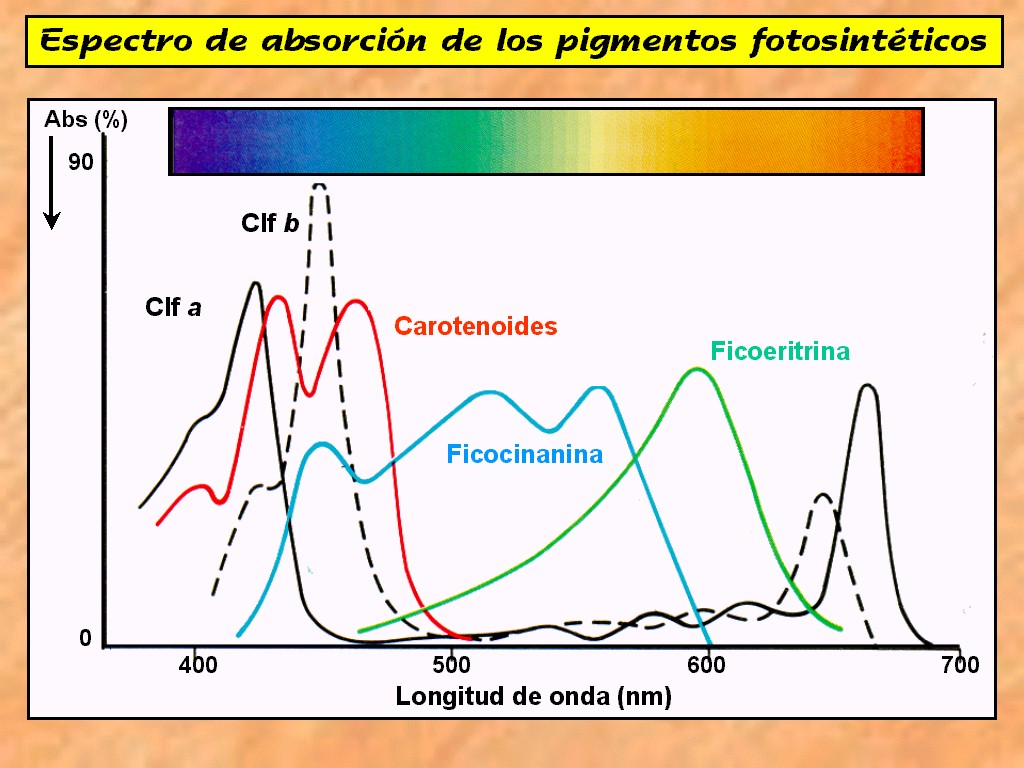
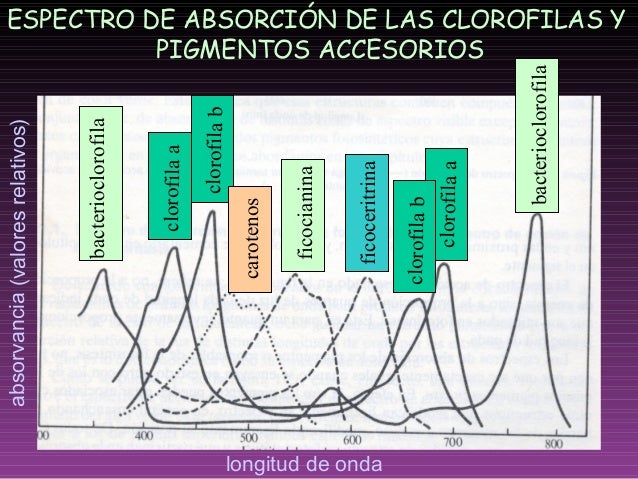
 Espectro de la luz solar.
Espectro de la luz solar.


 La luz, imprescindible para la vida vegetal.
La luz, imprescindible para la vida vegetal.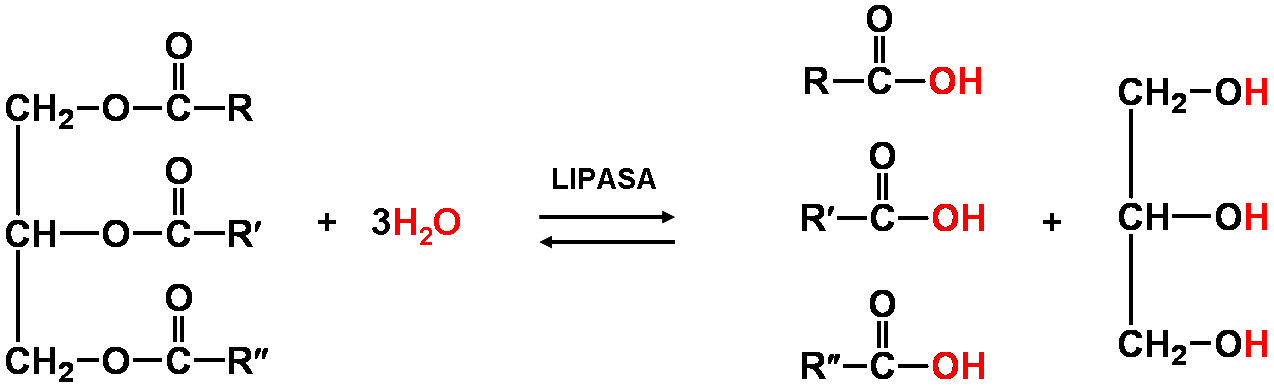
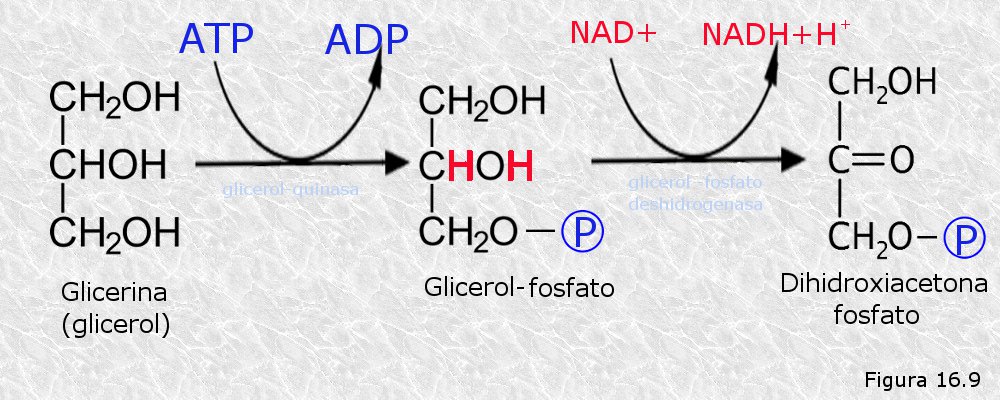
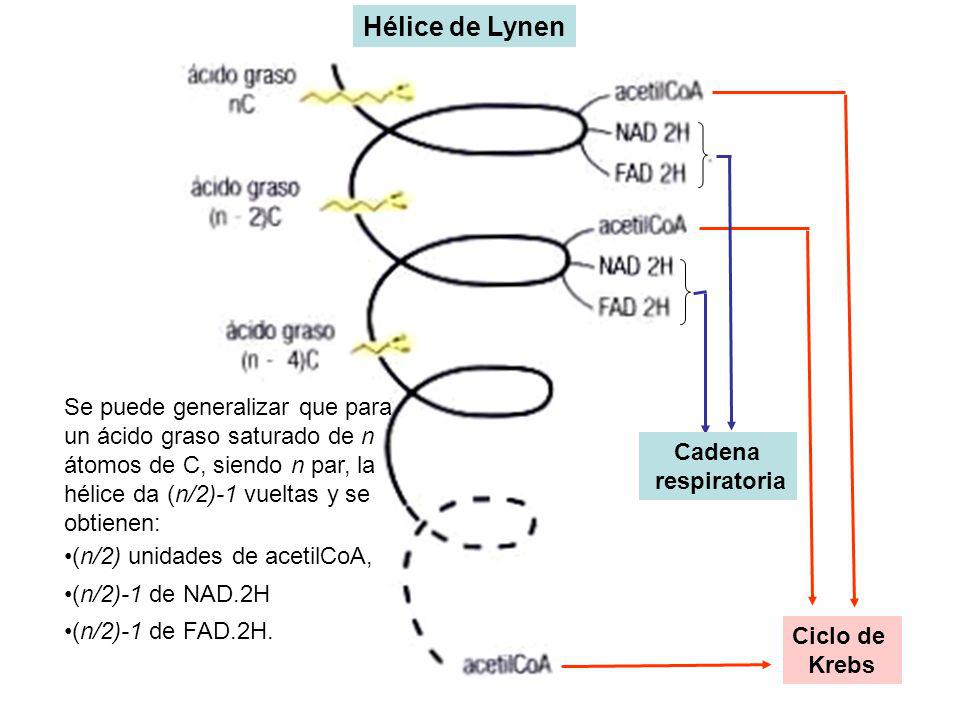



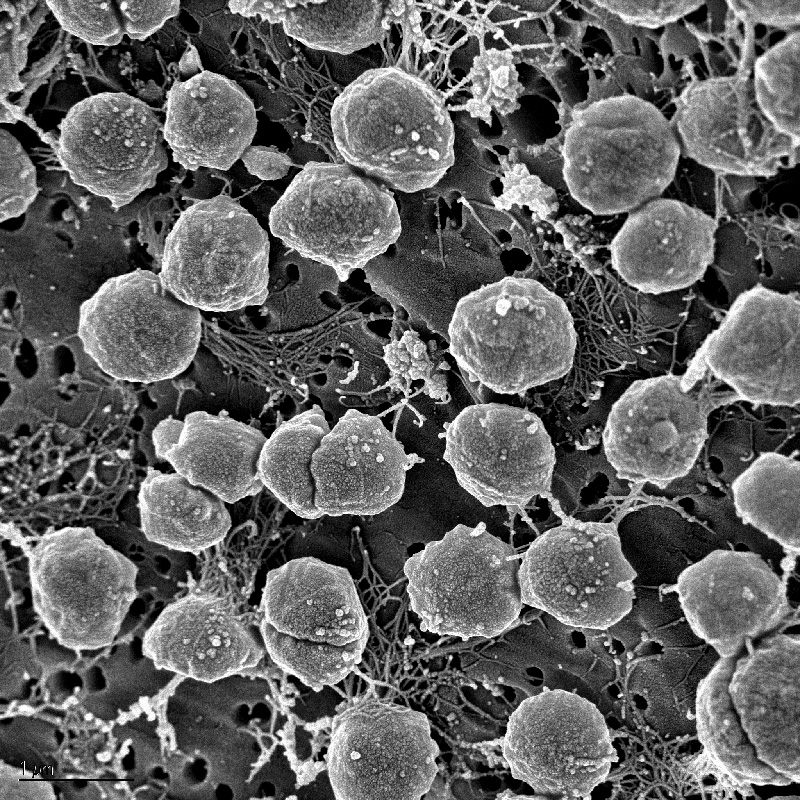

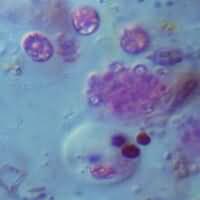 Bacterias púrpuras del azufre.
Bacterias púrpuras del azufre. Bacterias verdes del azufre.
Bacterias verdes del azufre.